トレーニング理論
筋トレの重量停滞を打破する『メカニカルストレス』の真実と密度
高重量至上主義による重量停滞や関節痛に悩む方へ。「高重量=筋肥大」の認知の歪みを解体し、時間変数を組み込んだV.P.M理論に基づく本質的なメカニカルストレスと「密度」の漸進法を解説します。
ブログを読み込み中...
高重量至上主義による重量停滞や関節痛に悩む方へ。「高重量=筋肥大」の認知の歪みを解体し、時間変数を組み込んだV.P.M理論に基づく本質的なメカニカルストレスと「密度」の漸進法を解説します。
筋肥大をストレス応答と捉え、時間変数を統合した密度指標V.P.MとV.P.Sを提唱。密度を目標ではなく適応の「結果」として観測し、精密なピーキングや初級者評価を可能にする、停滞打破のための新基準。
ベンチプレスの重量が 100kg でピタリと止まった。スクワットで限界を超えて追い込んでから、膝や腰の違和感が消えない。 「もっと重いものを扱わなければ筋肉は大きくならない」と信じ、関節の痛みを誤魔化しながらバーベルを握っていないだろうか。
重量の更新はモチベーションの源泉である。しかし、ある一定のラインから重量が伸び悩み、怪我のリスクばかりが跳ね上がる時期が必ず訪れる。もしあなたが今、そのような「頭打ち感」や慢性的な疲労に悩んでいるのなら、これまでのアプローチを根本から見直すタイミングかもしれない。
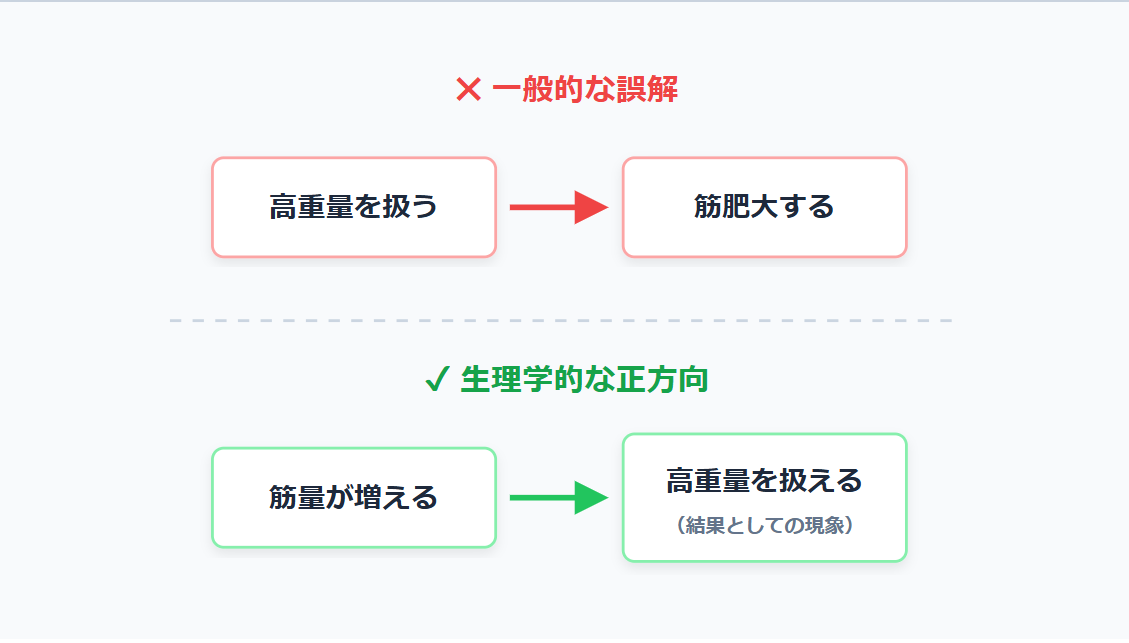
「高重量を使わないと筋量が増えない」
これは、トレーニング業界に深く根付いた認知の歪み(原因と結果の逆転)である。
生理学的な観点において、筋の断面積が大きい(筋量が多い)から高重量を扱えるのであって、高重量を振り回すこと自体が筋肥大の絶対条件ではない。重量や回数の漸進は、正しいトレーニングプロセスを経た「結果としての物理現象」に過ぎないのだ。
限界を超えて無理に重量を追うこと(チーティングや代償動作の誘発)は、中枢神経系を著しく抑制し、代謝的疲労を無駄に蓄積させる。筋肥大の本質から遠ざかるばかりか、関節へのダメージという負の遺産しか残らない。
従来の指導では、「強いメカニカルストレス」が「主観的な高重量の挙上」と誤って結びつけられてきた。
生理学およびバイオメカニクスの観点において、筋肥大のトリガーとなる本質的なメカニカルストレスとは——
「エキセントリック局面(伸張性収縮)における、筋長伸張に対する張力コントロール」
——を指す。
反動(SSC)を使って持ち上げたりした時点で、ターゲットとなるモーターユニット(運動単位)への負荷は分散する。結果として、筋肥大の司令塔である「mTORc1」へのシグナル伝達は抑制されてしまう。
重量(強度)や回数だけの漸進には限界がある。安全かつ持続的に成長を促すためには、絶対に切り離せない「時間」の概念を独立変数として取り入れた新基準——「密度」——による管理が必要だ。
「重量が停滞していても、密度(同時間内の仕事量)が向上していれば筋肥大の要件は満たされている」
この事実が、実践者から「常に重量を更新しなければならない」という心理的焦燥感を排除し、安全で持続可能な漸進を約束するのだ。
明日からのトレーニングでは、勇気を持って「引き算」を行ってほしい。
関節に頼った「静的支持」の重量設定を捨て、対象筋だけでコントロールできる「動的支持」の重量まで落とす。トップポジションで出力を切らず、エキセントリック局面で筋肉を「緩めながら」耐えることに集中する。
中枢神経系の抑制が働く前、つまり代償動作が出て V.P.S(動作の質)が低下する一歩手前でセットを終える。「限界まで追い込む」のではなく「質の高い動作を遂行する」ことに価値を置く。
トレーニングノートに重量と回数だけでなく、「所要時間」を記録する。次回以降、同じ重量・回数であっても、休憩時間をコントロールして少しでも時間が短縮されていれば(V.P.M が上がっていれば)、あなたの筋肉は確実に進化している。
「重さ」という物理的な呪縛から解放され、「密度」という生理学的な真実に向き合ったとき、あなたの体は再び成長の軌道に乗り始めるはずだ。

平木 翔(Sho Hiraki)
ふぃっとねす工房 代表パーソナルトレーナー / ストレングストレーナー
生理学とバイオメカニクスを基盤に、感覚に頼らない論理的なボディメイクを指導。自身も現役の競技者として「密度(V.P.M)」理論を体現し、その有効性を証明し続けている。
【実績・資格】